
* Back in June, I posted the first two parts of a three-part essay on Stephen Jay Gould and “punctuated equilibria” (PE). The idea was to commemorate the 50th anniversary of PE and the 20th anniversary of Gould’s death, both of which took place in 2022. You can find the original posts here and here. (And here is Part 3.)
Well, I’ve finally written the third part, which I’ll put up later this week. Before I do this, I thought I would share the first two parts again, this time in a single post. “Problematica” is written by Max Dresow…
PART 1: “How to change your life using punctuated equilibria”
The term “punctuated equilibria” (PE) was coined fifty years ago in a now-classic volume called Models in Paleobiology. Since then it has had an eventful career, first in paleontology, then in biology, and finally in sundry places like public policymaking and religious studies. Perhaps its most wayward application comes from an essay titled, “How to Change your Life Using Punctuated Equilibrium,” written by one of those life coach, business consultant types.* It won’t surprise you to learn that the author gets PE all wrong. To begin, he describes PE as “the way that most natural systems” evolve, which is either meaningless (since PE does not apply to most natural systems) or false. He contrasts PE with Darwinian gradualism, which is standard, but goes on to say that PE involves explosive blasts of evolution, which is incorrect. It soon becomes clear why he is talking about “explosions.” This is because he takes “[the] most well-known example of… punctuated equilibrium” to be the Cambrian explosion: the event in the fossil record corresponding to the emergence of abundant and diverse bilaterian animals. I once heard a similar suggestion on the BBC radio program In Our Time, but it’s dead wrong. The Cambridge explosion was a polyphyletic radiation; PE is a theory about the scaling of speciation in geological time. To call the former an example of the latter is to make a category mistake.
[* The terms “punctuated equilbria” and “punctuated equilibrium” are synonyms.]

The representation of punctuated equilibrium (right) versus gradual evolution (left) from the essay I mentioned above. Here, PE is misleadingly represented as taking place in a single unbranched lineage (this is sometimes called “punctuated anagenesis” to distinguish it from PE proper). A more standard representation of PE can be found here
I don’t intend these remarks to be patronizing, and for what it’s worth, I rather like the message the author spins out of his misunderstanding of PE. My point is simply this. “Punctuated equilibria” is one of the most misunderstood ideas in the paleontological literature. It is routinely mischaracterized, not just in popular writing but in scientific work as well. And much of the reason, I suspect, is that it is just so widely known. Lots of people have heard of PE, and almost as many have taken themselves to understand it, even without having read any of the primary literature. This is a recipe for misunderstanding.
But this essay is not about these relatively simple misunderstandings of PE. It is rather about a different kind of misunderstanding: one that concerns the history of the idea, particularly in relation to Stephen Jay Gould’s thinking about evolution. You may not have thought about this history before, but if you have you probably suspect that the publication of PE was the major event in Gould’s early career, overshadowing everything else. So it was, I suppose—but only in hindsight. Lots needed to happen before Gould could embrace PE as the “coordinating centerpiece” of his thought, as he put it in The Structure of Evolutionary Theory. Not least, he needed to renounce the idea that the history of major animal groups is a history of biological improvement, or “progress” in the sense of improved design. He also needed to repudiate his deep-seated preference for adaptive explanations in paleontology—but here I am getting ahead of myself.
This essay is split into three parts. You are reading the first part right now. The second and third parts will appear in the coming weeks. The remainder of Part 1 discusses what Gould was up to on the eve of “Punctuated equilibria: an alternative to phyletic gradualism” (the famous chapter in Models in Paleobiology, co-authored with Niles Eldredge). This included a promotional campaign for a vision of evolutionary paleontology that differed starkly from the one he would promote in his later work. Part 2 discusses PE itself, and argues that nothing in the original articulation of the theory forced Gould to abandon his vision for evolutionary paleontology or the associated ideas about evolution. But if nothing in PE forced Gould to abandon this vision for paleobiology, then why did he abandon it? And how did PE come to form the centerpiece of a radically different view of evolution associated with an almost diametrically opposed research program to the one he promoted in 1970? These are the questions I will take up in Part 3.
But first things first. What was Gould up to on the eve of “Punctuated Equilibria: An Alternative to Phyletic Gradualism”?
Portrait of the paleontologist as a young man
In 1966, a remarkable essay appeared in the journal Biological Reviews. Commissioned by Julian Huxley, it was written in a style that Huxley would have appreciated: not exactly lean, but witty and energetic in its own way. Its author was Stephen Jay Gould, and he began his review of allometry, or relative growth, by confessing “a nearly complete bias for seeking causes framed in terms of adaptation” (Gould 1966, 588). He then made clear that this extends even to changes that follow mechanically from increasing size: “for whatever reason complex organ systems may have originally developed, the expanded potential for further progress thus conferred… is a true and most significant phylogenetic advance” (591). Gould went on to observe that scientists “too often confine [their] explanatory attempts to immediate efficient causes,” perhaps as an inheritance “from the mechanical aspects of the Newtonian approach” (629). But “whereas the citation of efficient physiological mechanisms can explain the form of a wolf’s dentition, the response ‘all the better to eat you with my dear’ answers the question ‘why this specific form’ in another, and equally valid, sense.” Elsewhere, Gould states this position even more clearly. “[The] fundamental problem of evolutionary paleontology,” he writes, “[is] the explanation of form in terms of adaptation” (Gould 1967, 385).

Stephen Jay Gould at his typewriter
It must have surprised many readers to learn that this essay was penned by a graduate student, barely twenty-five years old. Gould had earned the commission by writing a fan letter of sorts to Huxley, who was by then an old man decades removed from his last scientific work of any note. Back in 1932, Huxley had kickstarted the study of allometry in his book, Problems of Relative Growth, which remains a seminal text. It is thus unsurprising to find Huxley’s influence radiating throughout the essay, just as it radiated through the whole of Gould’s early career.
Nowhere is this influence more evident than in Gould's remarks about progress, or as he sometimes says, “biological improvement.” Huxley was a man obsessed with progress. It was for him an idée fixe: nothing about evolution was more important than the “fact” that natural selection leads to improvement, including improvement of a general kind. Probably this is what made him such an effective avatar for the optimism that pervaded the postwar period. Huxley was the first Director-General of UNESCO, and sometimes wrote as if the purpose of evolution was to bring about the cosmopolitan liberalism of the 1950s. Yet for his younger acolyte, the importance of progress lay mostly in the promise it held for evolutionary paleontology.
Gould was not as intoxicated with progress as Huxley. He recognized, at least, that it was “a touchy subject,” since improvement “is so difficult to define and has so often been discussed with anthropocentric bias” (Gould 1968, 97). Still, it could hardly be dismissed on this count, since as he wrote in an important review,
The evolution of most major groups… is a history of mechanical improvement… [which] is recognized only when we can specify biological tasks, define the structures that fit them best and monitor the evolutionary changes that lead, usually in independent lineages, to new grades or functional… zones.” (Gould 1970, 111, emphasis added)
(“Grade” is a Huxleyan term meaning a group that occupies a shared level of morphological and physiological complexity, like the group of all reptiles excluding birds. The main difference between grades and “clades”— also a Huxleyan coinage— is that clades have a single evolutionary origin, whereas grades are presumed to have been attained by many independent lineages.)
But what does this mean for paleontology? First, some context. Gould was trained as a quantitative paleontologist during a time when there wasn’t much precedent for that sort of thing. His specialty was the analysis of form using multivariate statistics, which he applied to a variety of groups including land snails in the genera Poecilozonites and Cerion. He completed his PhD at Columbia University under Norman Newell, a leading invertebrate paleontologist, and John Imbrie, a pioneer of quantitative methods in paleontology who would go on to make important contributions to the theory of ice ages. Then it was on to Harvard University, where he would spend the remainder of his career as a professor of geology (and later zoology) and Curator of Invertebrate Paleontology at the Museum of Comparative Zoology.

Land snails in the genus Cerion, the subject of much of Gould’s empirical research after 1967
Gould arrived at Harvard in the fall of 1967. This was near the beginning of the episode retrospectively known as the “paleobiological revolution,” which saw the emergence of paleobiology as a distinct area of study focused on the quantitative analysis and interpretation of the history of life (Sepkoski 2012). The paleobiological revolution would come to be associated with the data-centric approach of Gould’s student, Jack Sepkoski, who compiled an exhaustive dataset of all the known marine fossil families and their stratigraphic ranges. But in 1967 Sepkoski was a college sophomore, and Gould had his own ideas about how to restore paleontology to a position of prominence among the evolutionary sciences. These were based on a coordination of multivariate biometry (or the statistical description of form), theoretical morphology (or the simulation of form in silico), and the experimental study of adaptation in fossils. Gould’s expertise was limited to statistical analysis; yet it was the third approach that formed the leading edge of the proposed research program. This is because he regarded it as the key to demonstrating the reality of mechanical improvement in large taxonomic groups, and it was this demonstration, he imagined, that would “reinstate paleontology as a source of new themes for evolutionary theory” (Gould 1970, 112).
The key was something called paradigm analysis. This was a technique that Martin Rudwick had developed for the inference of functions in fossils, and it went as follows. First, researchers would use their knowledge of contemporary organisms (along with the principles of engineering) to hypothesize about the possible functions of a fossil structure. Then they would build models for each hypothesis corresponding to the structure best able to perform the postulated function under the constraints imposed by the nature of the materials involved. These models he termed paradigms. After this they would run both the fossil (or a model of the fossilized structure) and the paradigms through experiments designed to test how well the fossil could have performed a function relative to the corresponding paradigm. The closer the fossil approximated the performance of the paradigm, the stronger was the case that the structure actually performed that function; although strictly speaking, all the analysis can show is that the fossil would have been capable of functioning in the manner indicated—it cannot show that it really did perform that function. (For more details on Rudwick and paradigm analysis, see the Appendix to Part 1 in the original version of this post.)

A paradigm for “zigzag deflections,” like those formed by the valve edges of certain brachiopods, on the assumption that their function is to filter out large food particles during filter feeding (see the Appendix for more details). The basic idea is that introducing zigzag deflections in an opening used for filter feeding increases the area of the slit without increasing the risk of harmful particles entering. From Rudwick (1964)
What interested Gould was the possibility that paradigm analysis could be performed longitudinally, on early and later members of a taxon, to test whether mechanical improvement had taken place in that taxon. This would involve, first, determining the likely function of a fossil structure, and then testing earlier and later instances of that structure to determine how closely each approximated a shared paradigm. Increases in efficiency would provide evidence that mechanical improvement had taken place in that taxon, and the convergence of many lineages upon a shared paradigm would provide evidence that the history of the taxon had seen the multiple evolution of a mechanically optimal structure.
Why did such evidence matter? Paleontologists had long noted that convergent evolution is a ubiquitous feature of the fossil record, with one even calling its recognition “the most distinctive contribution that paleontology has made to the modern evolutionary synthesis” (Sylvester-Bradley 1959, 193). It was also widely held that many groups of animals were “grades of improvement attained in the same way by many [separate] lineages” (Gould 1976, 119). (Examples included reptiles, mammals, and even metazoans as a whole.) What Gould hoped to show was that widespread parallelism and convergence could be ascribed to “the evolution, in many lineages, of features that improve the basic design of [a] taxon” (Gould 1970, 110). This would advance evolutionary theory by demonstrating what previous workers had only suggested: that a major pattern in life’s history resulted from natural selection shaping many lineages to perform shared functions. It would also support the notion that major animal groups are “grades of improvement” achieved through polyphyletic transitions to better functional arrangements: more efficient jaw mechanisms in fishes, say. And because the evidence would come from fossils, the contribution would be a distinctively paleontological one—just what was needed to pull paleontology out of the doldrums and establish it as an active and vital contributor to evolutionary science.
Huxley would have been proud. In the event, though, the research program failed to materialize. I know of just one attempt to apply paradigm analysis in the manner Gould indicated, by Martin Rudwick and his student Richard Cowen (see below). Later paleontologists seem hardly to have noticed the suggestion, or if they noticed it, they did not pursue it.
As for Gould, he was soon busying himself with other attempts to remediate paleontology’s low standing: first stochastic models of clade dynamics, then a framework for interpreting developmental changes in adaptive terms, and finally, a “hierarchical expansion” of evolutionary theory with PE as its fulcrum (Sepkoski 2012; Dresow 2019). There is much that could be said about this frenzied period in Gould’s career, and I will say some of it in Part 3 of this essay. For now, the thing to notice is that PE only assumed a central place in his thought near the end of the 1970s, not in 1972, when “Punctuated Equilibria: An Alternative to Phyletic Gradualism” appeared. Why was this, and why did Gould continue to defend a basically adaptationist and progressivist view of evolution into the second half of the decade? These are questions I take up in Part 2.
Part 2: “PARADOX OF STASIS”
In The Structure of Evolutionary Theory, Stephen Jay Gould devotes about 300 pages to punctuated equilibria (PE), spanning a single sprawling chapter. He justifies the indulgence by claiming that PE permits species-level selection by individuating species in geological time. So, PE is foundational to the hierarchical expansion of evolutionary theory that is the book’s main theme. But this does not exhaust PE’s importance for Gould, since the idea also functions as a “coordinating centerpiece… for a larger and coherent set of mostly iconoclastic concerns [in evolutionary theory and the history of life]” (Gould 2002, 37). One of these is a critique of “Darwinian functionalism,” which amounts to a criticism of the idea that all evolutionary phenomena flow from the operation of natural selection in local populations. Another is an assault on the notion that progress supplies an important vector in the history of life. (Those of you who have read the first part of this essay will have noticed that these themes represent near-perfect inversions of the commitments Gould held as a young researcher.)

The Structure of Evolutionary Theory in all its heft, with punctuated equilibria as its “centerpiece”
The puzzle I will try to solve in the third and final part of this essay is how PE came to play this coordinating role, given that Gould remained committed to adaptationism and a broadly progressive view of life’s history even after the publication of “Punctuated equilibria: an alternative to phyletic gradualism” (Eldredge and Gould 1972). Yet before we come to this we must ask another question: why did the publication of PE in 1972 fail to rouse Gould from his dogmatic slumber? If PE was pregnant with major implications for evolutionary theory, why did Eldredge and Gould fail to bring them out? In short, how did the guy described in Part 1 of this essay weather the publication of PE with his basic view of evolution mostly intact?
The answer is that nothing in the original articulation of PE forced Gould to abandon either his adaptationism or his confidence that progress supplies an overarching theme in the history of life. So he didn’t—at least not right away. His views would shift considerably in the mid-1970s, and by 1980 would resemble those expressed in Structure. But that leaves the first half of the decade to make sense of. This was a period of relative stasis, at least as regards Gould’s basic ideas about evolution, which was terminated by an abrupt shift. The present part of this essay seeks to come to grips with the stasis; Part 3 concerns the punctuation.
Because David Sepkoski has already described the origin of PE in Rereading the Fossil Record (Chapter 5), I will mostly ignore this history here. Instead, I am going to focus on Eldredge and Gould’s discussion of trends, which they would later advance as the basis of a new view of macroevolution compelling a hierarchical revision of evolutionary theory. What I want to show is the surprising extent to which this treatment meshed with Gould’s other ideas about evolution, in particular, his commitment to the importance of biological improvement driven by natural selection.*
[* The section of “Punctuated equilibria” dealing with trends was written by Eldredge, but this is irrelevant to my argument. What matters is that Gould signed off on it, and that this involved no reworking of his basic ideas about evolution or the goals of evolutionary paleontology.]
Punctuated equilibria
To begin, a primer. PE is based on a model of speciation developed by Ernst Mayr, in which a (typically small) population becomes isolated from a source population at the periphery of its range. After a period of isolation the “peripheral isolate” loses its ability to interbreed with the source population, at which point it graduates to the rank of a new species. Mayr proposed that these isolates are good candidates to undergo rapid evolution owing to their small size and likely habitation in new environments. They are also more likely than source populations to incubate evolutionary novelties, including entirely new structures and ways of life.

Ernst Mayr, right, outside his new office in the Museum of Comparative Zoology. The man on the left is the renowned paleontologist and outgoing director of the MCZ, Alfred Romer
This is why. During the normal run of a species’ existence, its genes form a tightly coadapted bundle, precluding extensive reworking of the gene pool. During speciation, by contrast, a reduction in population size exposes homozygotes to selection and more generally “produce[s] a sudden change of the genetic environment of most loci” (Mayr 1954, 170). At this point all hell breaks loose, genetically speaking, and this permits the population to “pass from one well integrated and rather conservative condition… to another” (172). It is the genetic equivalent of moving from Louis XVI to Napoleon via the Terror. It is how biological Ancien Régimes fall, and are replaced by newer and, perhaps, better ones.
What Eldredge and Gould asked was what we should expect to see in sedimentary successions if this model is correct. Their answer is that we should expect a pattern of morphological stasis (corresponding to the stratigraphic range of the source population) punctuated by rapid change (corresponding to the reinvasion of a formerly isolated peripheral offshoot, say). This will generate an apparently gappy record lacking smooth morphological transitions. But if the model is correct, these gaps are artifacts of the speciation process itself. The intermediate forms exist(ed), they just don’t exist where a naive observer would expect to find them (and perhaps because of the small size of peripheral isolates they are less likely to be preserved as fossils).* By contrast, the morphological stability of the source population is not an artifact. Instead, it is evidence that something was preventing the population from changing: in Mayr’s terminology, the “homeostasis” of coadapted gene pools.
[* Mayr himself makes this point in his (1954): “It seems to me that many puzzling phenomena, particularly those that concern paleontologists, are elucidated by a consideration of [peripherally isolated] populations. This concerns primarily the phenomena of unequal (and particularly rapid) evolutionary rates, breaks in evolutionary sequences and apparent saltations, and finally the origin of new ‘types’” (175).]
So far there is nothing in this treatment to shake the foundations of evolutionary theory. As one reviewer of Models in Paleobiology put it, the proposal “will sound novel only to earth scientists who have not read anything in 20 years[!]” (This is somewhat uncharitable since Eldredge and Gould make much of the ubiquity of morphological stasis, and this is not cribbed from Mayr.) But Eldredge and Gould stake a claim to novelty in their discussion of large-scale evolutionary trends. They begin with a puzzle. Grant that species spend the majority of their histories in morphological stasis. How then are paleontologists to explain the existence of long-term directional changes in higher taxa? Slow and steady evolution in lineages will not do the trick, since on this view most change is cramped into relatively infrequent speciation events separating a whole lotta nothing. Perhaps natural selection acts to shift morphology in the same direction whenever new species happen to arise, but this seems improbable. Instead, Eldredge and Gould
envision multiple ‘explorations’ or ‘experimentations’ (see Schaeffer 1965)—i.e., invasions, on a stochastic basis, of new environments by peripheral isolates. There is nothing inherently directional about these invasions. However, a subset of these new environments might, in the context of inherited [sic] genetic constitution in the ancestral components of the lineage, lead to a new and improved efficiency. Improvement would be consistently greater within this hypothetical subset of local conditions that a population might invade. The overall effect would then be one of net, apparently directional change: but as with the case of selection upon mutations, the initial variation would be stochastic with respect to this change.
Basically, if speciation is random with respect to the direction of an ongoing trend, and if there is some reason that species varying in the direction of the trend are likely to do better than those varying in another direction, then a process of differential survival among incipient species will drive the trend forward, generating directional change.
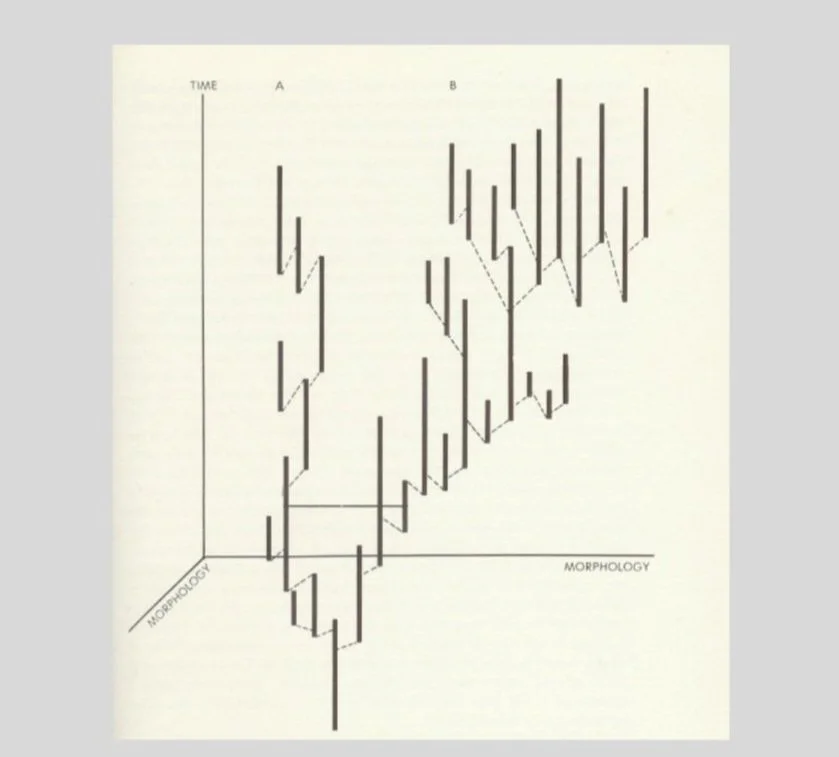
Eldredge and Gould’s illustration of a directional trend (B), with morphology pictured on the horizontal axis and time on the vertical. Bold vertical lines represent morphologically stable species; dashed lines are speciation events. As Eldredge and Gould write: “Though a retrospective pattern of directional selection might be fitted as a straight line in (B), the actual pattern is stasis within species, and differential success of species exhibiting morphological change in a particular direction”
If the expression “new and improved efficiency” made your ears tingle, then I’ve done my job. As you may have guessed, I’m going to make a big deal out of this in the next section. First, though, I want to glance ahead to 1977, when the sequel to “Punctuated Equilibria: An Alternative to Phyletic Gradualism” appeared. Titled “Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered,” it presents the same basic story about trends, with random speciation (analogous to mutation) followed by the non-random success of a subset of speciations (natural selection). There is one big difference, however. In the new paper, the emphasis falls on the “decoupling” of micro- and macroevolution via punctuated equilibria, and its implication for the explanation of general patterns in the history of life. In this formulation, “speciation interposes itself as an intermediate level between macroevolutionary trends and evolutionary events within populations.” It follows, Gould and Eldredge claim, that the explanation of macroevolutionary trends must involve processes other than those active in local populations: in particular, processes that operate on populations of species.
A full discussion of this reimagining of PE must await the third part of this essay. The point of raising it here is simply to observe that claims of “decoupling” are not simply a restatement of the view given in Eldredge and Gould (1972). The idea that macroevolution is decoupled from microevolution, and that major features in the history of life owe to an irreducible domain of macroevolutionary causes, was an addition to what Eldredge had written in 1972. Perhaps it was implicit in the original treatment, but anyway it was not highlighted, and probably it was not even recognized. I suspect it was not recognized because in order to embrace decoupling, Eldredge and especially Gould had to abandon certain ideas that they were not keen to give up on in 1972. This took an external stimulus. Regardless, the point for now is that nothing in the logic of PE itself forced Eldredge or Gould to rethink the centrality of adaptation and progress in the history of life. So they didn't, until they had reason to.
Punctuation and progress
To see why this is the case, we need to return to their original discussion of evolutionary trends.
Recall that for Eldredge and Gould, speciation involves the adaptation of an isolated segment of a population to novel environmental conditions. By itself, this is not expected to produce directional trends in higher taxa. But imagine that some isolates find themselves in environments to which they are especially well-fitted, despite having evolved to live in rather different environments (the phenomenon of preadaptation). Then we can expect, as George Simpson puts it in Tempo and Mode in Evolution, that “[such] groups [will be] subjected to strong selection pressure and [will] evolve rapidly in the further direction of adaptation to the new [conditions]” (123). What Eldredge and Gould propose is that this process will sometimes produce “net, apparently directional change” in a group of lineages. But when, exactly? In their words: when a subset of environments exist whose invasion yields “new and improved efficiency,” permitting certain isolates to survive when so many others perish.
In all this adaptation is the key consideration. But not all adaptations are created equal. According to an old paleontological adage, adaptive specialization is the enemy of evolutionary progress. On this view, specialized lineages sacrifice evolutionary plasticity for short term adaptive gains. The result is the improvement of some organs at the expense of others, leading to what Julian Huxley calls “limiting” or “one-sided” improvement (Huxley 1942, 564). (Think of a parasite, which becomes better at its craft by shedding certain organs.) However, there also exist adaptations that contribute to “raising [the] upper level of all-round functional efficiency.” These adaptations are the stuff of open-ended evolutionary progress. They are the source, ultimately, of higher taxa representing new grades of functional organization. In a word, they are exactly the kind of adaptations that are likely to be incorporated into large-scale evolutionary trends.
Are these non-limiting adaptations what Eldredge and Gould have in mind when they speak about “new and improved efficiency”? There are at least two reasons to think so. First, Eldredge and Gould cite Bobb Schaeffer in connection with their concept of “‘explorations’ or ‘experimentations’ (see Schaeffer 1965)—i.e., invasions, on a stochastic basis, of new environments by peripheral isolates.” This matters because Schaeffer’s 1965 paper is about “[the] role of experimentation in the origin of higher levels of organization,” which Schaeffer understands, following Huxley, as grades of increasing functional efficiency. Schaeffer was a paleontologist at the American Museum and a professor at Columbia University, where Eldredge and Gould were graduate students in the 1960s. They must have known that his concept of experimentation involves the “trying out” of sets of character until (so to speak) one of them achieves an overall improvement. As Schaeffer writes, “Each successful [character] combination must be adapted to a particular ecological niche, but some combinations may also represent improved solutions to functional problems common to many different niches.” Not all adaptations are created equal.*
[* We should be wary of over-interpreting a single citation, but if the analogy of speciation to “experimentation” is taken at all seriously, then successful “experiments” will be those that experience a broad and non-limiting kind of adaptive improvement.]

Schaeffer’s (1965) diagram depicting “the Phylogeny of Amphibia in relation to levels of organization.” The numbers, (1)–(3), refer to successful “experiments” in the “amphibian direction.” Each corresponds to a “broad adaptation” representing the best possible solution to a basic functional problem. Crosses represent failed experiments, or lineages that failed to achieve the new organizational grade
The second reason is a conceptual one. If all speciation involves adaptation to local conditions, then Eldredge and Gould cannot specify the subset of a directional trend by gesturing at adaptation or “improvement” as such. They must have a special kind of improvement in mind, like an increase in overall efficiency as opposed to the refinement of a narrow functional capacity. And as luck would have it, there is some textual evidence that this is the case. In a book chapter published in 1977, Gould remarks that many nineteenth century paleontologists rejected natural selection because it offered no toehold for a belief in progress (a dubious historical claim on Gould’s part). These paleontologists were right, Gould claims: natural selection operating within populations generates no overall directionality, only a toing and froing of population in response to shifting local exigencies. However, “once we discard the shackles of phyletic gradualism as an explanation for ‘trends’, we can see that the operation of natural selection in evolutionary time can yield direction” (Gould 1977, 22). His explanation follows Eldredge and Gould (1972), but is more explicit at key points:
The primary events of speciation yield no direction, for they only adapt populations to local environments. But all speciations do not have an equal phyletic longevity or an equal opportunity for further speciation. Trends represent the differential success of subsets from a random spectrum of speciations. Improved biomechanical efficiency, for example, represents one pathway to adaptation in local environments. The species that follow this path—rather than the acquisition of a limiting, morphological specialization—might form the subset of a directional trend. (Gould 1977, 22, emphases added)
Admittedly, Gould writes “for example,” which indicates that improved biomechanical efficiency isn’t the only way species become incorporated into a trend. But that isn’t the point. The point is that this rendering of PE “saves the phenomenon” at the center of Gould’s early vision for evolutionary paleontology—improvement in the basic design of a large taxon. And this supplies a satisfying answer to the question posed above: how did Gould weather the publication of PE with his basic view of evolution mostly intact?
* * *
I titled this essay “Paradox of Stasis” because there is something superficially paradoxical about the stability of Gould’s thinking between 1970 and 1975.* A naïve observer, confronted with evidence of Gould’s adaptationism and progressivism, might understandably look to PE as a kind of heel turn. Sure, before PE Gould said some pretty un-Gould-y things about evolution. But after 1972 things must have clicked into place. —Right?
Wrong. PE did not mark a sea change in Gould’s thought, despite the important role it would come to play in his mature view of life. The reason is that PE, and especially the critical section on trends, was entirely compatible with his youthful view of evolution. It was only after 1975 that the tide began to shift for various reasons to be explored in the next and final part of this essay. As a result, PE would be thrust into the center of Gould’s renewed campaign to establish paleontology as an essential and irreducible contributor to evolutionary theory.
[* The expression “paradox of stasis” also has a meaning in the paleontological literature. Here is a nice philosophical treatment by Jonathan Kaplan, and check out this old post by Derek Turner.]
References
Dresow, M. 2017. Before hierarchy: the rise and fall of Stephen Jay Gould’s first macroevolutionary synthesis. History and Philosophy of the Life Sciences 39:1–30.
Dresow, M. 2019. Macroevolution evolving: punctuated equilibria and the roots of Stephen Jay Gould’s second macroevolutionary synthesis. Studies in History and Philosophy of Biological and Biomedical Science, 75:15–23.
Dresow, M. 2019. Gould’s laws: a second perspective. Biology & Philosophy, https://doi.org/10.1007/s10539-019-9698-7.
Eldredge, N. and Gould, S.J. 1972. Punctuated equilibria: an alternative to phyletic gradualism. In T.J.M. Schopf (Ed.), Models in Paleobiology, pp. 82–115. San Francisco: Cooper & Co.
Gould, S.J. 1966. Allometry and size in ontogeny and phylogeny. Biological Reviews 41:587–640.
Gould, S.J. 1967. Evolutionary patterns in Pelycosaurian reptiles: a factor-analytic study. Evolution 21:385–401.
Gould, S.J. 1968. Ontogeny and the explanation of form: an allometric analysis. Memoir (The Paleontological Society), Vol. 2, Supplement to Volume 42 of the Journal of Paleontology, pp. 81–91.
Gould, S.J. 1970. Evolutionary paleontology and the science of form. Earth-Science Reviews 6:77–119.
Gould, S.J. 1976. Grades and clades revisited. In R.B. Masterton, W. Hodos and H. Jerison (Eds.), Evolution, Brain, and Behaviour, pp. 115–122. Hillsdale: Lawrence Erlbaum Associates.
Gould, S.J. 1977. Eternal metaphors in paleontology. In A. Hallam (Ed.). Patterns of Evolution as Illustrated by the Fossil Record, pp. 1–26. Amsterdam: Elsevier.
Gould, S.J. 2002. The Structure of Evolutionary Theory. Cambridge, MA: Belknap Press.
Gould, S.J. and Eldredge, N. 1977. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology 3:115–151.
Huxley, J. 1932. Problems of Relative Growth. New York: The Dial Press.
Huxley, J. 1942. Evolution: The Modern Synthesis. New York: John Wiley & Sons.
Mayr, E. 1954. Change of genetic environment and evolution. In J. Huxley, A.C. Hardy, E.B. Ford (Eds.), Evolution as a Process, pp. 157–180. London: Gorge Allen & Unwin Ltd.
Rudwick, M.J.S. 1964. The inference of function from structure in fossils. The British Journal for the Philosophy of Science 7:27–40.
Schaeffer, B. 1965. The role of experimentation in the origin of higher levels of organization. Systematic Zoology 14:318–336.
Sepkoski, D. 2012. Rereading the Fossil Record: The Growth of Paleobiology as an Evolutionary Discipline. Chicago: University of Chicago Press.
Simpson, G.G. 1944. Tempo and Mode in Evolution. New York: Columbia University Press.