
* In this three-part installment of “Problematica,” Max Dresow explores the history of punctuated equilibria, which celebrated its fiftieth anniversary last year. This is Part 1. Part 2 and Part 3 are also available to read…
The term “punctuated equilibria” (PE) was coined fifty years ago in a now-classic volume called Models in Paleobiology. Since then it has had an eventful career, first in paleontology, then in biology, and finally in sundry places like public policymaking and religious studies. Perhaps its most wayward application comes from an essay titled, “How to Change your Life Using Punctuated Equilibrium,” written by one of those life coach, business consultant types.* It won’t surprise you to learn that the author gets PE all wrong. To begin, he describes PE as “the way that most natural systems” evolve, which is either meaningless (since PE does not apply to most natural systems) or false. He contrasts PE with Darwinian gradualism, which is standard, but goes on to say that PE involves explosive blasts of evolution, which is incorrect. It soon becomes clear why he is talking about “explosions.” This is because he takes “[the] most well-known example of… punctuated equilibrium” to be the Cambrian explosion: the event in the fossil record corresponding to the emergence of abundant and diverse bilaterian animals. I once heard a similar suggestion on the BBC radio program In Our Time, but it’s dead wrong. The Cambridge explosion was a polyphyletic radiation; PE is a theory about the scaling of speciation in geological time. To call the former an example of the latter is to make a category mistake.
[* The terms “punctuated equilibria” and “punctuated equilibrium” are synonyms.]

The representation of punctuated equilibrium (right) versus gradual evolution (left) from the essay I mentioned above. Here, PE is misleadingly represented as taking place in a single unbranched lineage (this is sometimes called “punctuated anagenesis” to distinguish it from PE proper). A more standard representation of PE can be found here
I don’t intend these remarks to be patronizing, and for what it’s worth, I rather like the message the author spins out of his misunderstanding of PE. My point is simply this. “Punctuated equilibria” is one of the most misunderstood ideas in the paleontological literature. It is routinely mischaracterized, not just in popular writing but in scientific work as well. And much of the reason, I suspect, is that it is just so widely known. Lots of people have heard of PE, and almost as many have taken themselves to understand it, even without having read any of the primary literature. This is a recipe for misunderstanding.
But this essay is not about these relatively simple misunderstandings of PE. It is rather about a different kind of misunderstanding: one that concerns the history of the idea, particularly in relation to Stephen Jay Gould’s thinking about evolution. You may not have thought about this history before, but if you have you probably suspect that the publication of PE was the major event in Gould’s early career, overshadowing everything else. So it was, I suppose—but only in hindsight. Lots needed to happen before Gould could embrace PE as the “coordinating centerpiece” of his thought, as he put it in The Structure of Evolutionary Theory. Not least, he needed to renounce the idea that the history of most animal groups is a history of biological improvement, or “progress” in the sense of increased mechanical efficiency. He also needed to repudiate his deep-seated preference for adaptive explanations in paleontology— but here I am getting ahead of myself.
This essay is split into three parts. You are reading the first part right now. The second and third parts will appear in the coming weeks. The remainder of Part 1 discusses what Gould was up to on the eve of “Punctuated equilibria: an alternative to phyletic gradualism” (the famous chapter in Models in Paleobiology, co-authored with Niles Eldredge). This included a promotional campaign for a vision of evolutionary paleontology that differed starkly from the one he would promote in his later work. Part 2 discusses PE itself, and argues that nothing in the original articulation of the theory forced Gould to abandon his vision for evolutionary paleontology or the associated ideas about evolution. But if nothing in PE forced Gould to abandon this vision for paleobiology, then why did he abandon it? And how did PE come to form the centerpiece of a radically different view of evolution associated with an almost diametrically opposed research program to the one he promoted in 1970? These are the questions I will take up in Part 3.
But first things first. What was Gould up to on the eve of “Punctuated Equilibria: An Alternative to Phyletic Gradualism”?
Portrait of the paleontologist as a young man
In 1966, a remarkable essay appeared in the journal Biological Reviews. Commissioned by Julian Huxley, it was written in a style that Huxley would have appreciated: not exactly lean, but witty and energetic in its own way. Its author was Stephen Jay Gould, and he began his review of allometry, or relative growth, by confessing “a nearly complete bias for seeking causes framed in terms of adaptation” (Gould 1966, 588). He then made clear that this extends even to changes that follow mechanically from increasing size: “for whatever reason complex organ systems may have originally developed, the expanded potential for further progress thus conferred… is a true and most significant phylogenetic advance” (591). Gould went on to observe that scientists “too often confine [their] explanatory attempts to immediate efficient causes,” perhaps as an inheritance “from the mechanical aspects of the Newtonian approach” (629). But “whereas the citation of efficient physiological mechanisms can explain the form of a wolf’s dentition, the response ‘all the better to eat you with my dear’ answers the question ‘why this specific form’ in another, and equally valid, sense.” Elsewhere, Gould states this position even more clearly. “[The] fundamental problem of evolutionary paleontology,” he writes, “[is] the explanation of form in terms of adaptation” (Gould 1967, 385).

Stephen Jay Gould at his typewriter
It must have surprised many readers to learn that this essay was penned by a graduate student, barely twenty-five years old. Gould had earned the commission by writing a fan letter of sorts to Huxley, who was by then an old man decades removed from his last scientific work of any note. Back in 1932, Huxley had kickstarted the study of allometry in his book, Problems of Relative Growth, which remains a seminal text. It is thus unsurprising to find Huxley’s influence radiating throughout the essay, just as it radiated through the whole of Gould’s early career.
Nowhere is this influence more evident than in Gould's remarks about progress, or as he sometimes says, “biological improvement.” Huxley was a man obsessed with progress. It was for him an idée fixe: nothing about evolution was more important than the “fact” that natural selection leads to improvement, including improvement of a general kind. Probably this is what made him such an effective avatar for the optimism that pervaded the postwar period. Huxley was the first Director-General of UNESCO, and sometimes wrote as if the purpose of evolution was to bring about the cosmopolitan liberalism of the 1950s. Yet for his younger acolyte, the importance of progress lay mostly in the promise it held for evolutionary paleontology.
Gould was not as intoxicated with progress as Huxley. He recognized, at least, that it was “a touchy subject,” since improvement “is so difficult to define and has so often been discussed with anthropocentric bias” (Gould 1968, 97). Still, it could hardly be dismissed on this count, since as he wrote in an important review,
The evolution of most major groups… is a history of mechanical improvement… [which] is recognized only when we can specify biological tasks, define the structures that fit them best and monitor the evolutionary changes that lead, usually in independent lineages, to new grades or functional… zones.” (Gould 1970, 111, emphasis added)
(“Grade” is a Huxleyan term meaning a group that occupies a shared level of morphological and physiological complexity, like the group of all reptiles excluding birds. The main difference between grades and “clades”— also a Huxleyan coinage— is that clades have a single evolutionary origin, whereas grades are presumed to have been attained by many independent lineages.)
But what does this mean for paleontology? First, some context. Gould was trained as a quantitative paleontologist during a time when there wasn’t much precedent for that sort of thing. His specialty was the analysis of form using multivariate statistics, which he applied to a variety of groups including land snails in the genera Poecilozonites and Cerion. He completed his PhD at Columbia University under Norman Newell, a leading invertebrate paleontologist, and John Imbrie, a pioneer of quantitative methods in paleontology who would go on to make important contributions to the theory of ice ages. Then it was on to Harvard University, where he would spend the remainder of his career as a professor of geology (and later zoology) and Curator of Invertebrate Paleontology at the Museum of Comparative Zoology.

Land snails in the genus Cerion, the subject of much of Gould’s empirical research after 1967
Gould arrived at Harvard in the fall of 1967. This was near the beginning of the episode retrospectively known as the “paleobiological revolution,” which saw the emergence of paleobiology as a distinct area of study focused on the quantitative analysis and interpretation of the history of life (Sepkoski 2012). The paleobiological revolution would come to be associated with the data-centric approach of Gould’s student, Jack Sepkoski, who compiled an exhaustive dataset of all the known marine fossil families and their stratigraphic ranges. But in 1967 Sepkoski was a college sophomore, and Gould had his own ideas about how to restore paleontology to a position of prominence among the evolutionary sciences. These were based on a coordination of multivariate biometry (or the statistical description of form), theoretical morphology (or the simulation of form in silico), and the experimental study of adaptation in fossils. Gould’s expertise was limited to statistical analysis; yet it was the third approach that formed the leading edge of the proposed research program. This is because he regarded it as the key to demonstrating the reality of mechanical improvement in large taxonomic groups, and it was this demonstration, he imagined, that would “reinstate paleontology as a source of new themes for evolutionary theory” (Gould 1970, 112).
The key was something called paradigm analysis. This was a technique that Martin Rudwick had developed for the inference of functions in fossils, and it went as follows. First, researchers would use their knowledge of contemporary organisms (along with the principles of engineering) to hypothesize about the possible functions of a fossil structure. Then they would build models for each hypothesis corresponding to the structure best able to perform the postulated function under the constraints imposed by the nature of the materials involved. These models he termed paradigms. After this they would run both the fossil (or a model of the fossilized structure) and the paradigms through experiments designed to test how well the fossil could have performed a function relative to the corresponding paradigm. The closer the fossil approximated the performance of the paradigm, the stronger was the case that the structure actually performed that function; although strictly speaking, all the analysis can show is that the fossil would have been capable of functioning in the manner indicated—it cannot show that it really did perform that function. (For more details on Rudwick and paradigm analysis, see the Appendix, below.)

A paradigm for “zigzag deflections,” like those formed by the valve edges of certain brachiopods, on the assumption that their function is to filter out large food particles during filter feeding (see the Appendix for more details). The basic idea is that introducing zigzag deflections in an opening used for filter feeding increases the area of the slit without increasing the risk of harmful particles entering. From Rudwick (1964)
What interested Gould was the possibility that paradigm analysis could be performed longitudinally, on early and later members of a taxon, to test whether mechanical improvement had taken place in that taxon. This would involve, first, determining the likely function of a fossil structure, and then testing earlier and later instances of that structure to determine how closely each approximated a shared paradigm. Increases in efficiency would provide evidence that mechanical improvement had taken place in that taxon, and the convergence of many lineages upon a shared paradigm would provide evidence that the history of the taxon had seen the multiple evolution of a mechanically optimal structure.
Why did such evidence matter? Paleontologists had long noted that convergent evolution is a ubiquitous feature of the fossil record, with one even calling its recognition “the most distinctive contribution that paleontology has made to the modern evolutionary synthesis” (Sylvester-Bradley 1959, 193). It was also widely suspected that many groups of animals were “grades of improvement attained in the same way by many [separate] lineages” (Gould 1976, 119). (Examples included reptiles, mammals, and even metazoans as a whole.) What Gould hoped to show was that widespread parallelism and convergence could be ascribed to “the evolution, in many lineages, of features that improve the basic design of [a] taxon” (Gould 1970, 110). This would advance evolutionary theory by demonstrating what previous workers had only suggested: that a major pattern in life’s history resulted from natural selection shaping many lineages to perform shared functions. It would also support the notion that major animal groups are “grades of improvement” achieved through polyphyletic transitions to better functional arrangements: more efficient jaw mechanisms in fishes, say. And because the evidence would come from fossils, the contribution would be a distinctively paleontological one—just what was needed to pull paleontology out of the doldrums and establish it as an active and vital contributor to evolutionary science.
Huxley would have been proud. In the event, though, the research program failed to materialize. I know of just one attempt to apply paradigm analysis in the manner Gould indicated, by Martin Rudwick and his student Richard Cowen (see below). Later paleontologists seem hardly to have noticed the suggestion, or if they noticed it, they did not pursue it.
As for Gould, he was soon busying himself with other attempts to remediate paleontology’s low standing: first stochastic models of clade dynamics, then a framework for interpreting developmental changes in adaptive terms, and finally, a “hierarchical expansion” of evolutionary theory with PE as its fulcrum (Sepkoski 2012; Dresow 2019). There is much that could be said about this frenzied period in Gould’s career, and I will say some of it in Part 3 of this essay. For now, the thing to notice is that PE only assumed a central place in his thought near the end of the 1970s, not in 1972, when “Punctuated Equilibria: An Alternative to Phyletic Gradualism” appeared. Why was this, and why did Gould continue to defend a basically adaptationist and progressivist view of evolution into the second half of the decade? These are questions I take up in Part 2.
Appendix: Martin Rudwick and “the method of paradigms”
The problem of functional inference has long struck paleontologists as a hairy one, especially when a structure has no close analog in the living biota. Invertebrate paleontologists were particularly slow to apply functional reasoning to fossils, partly owing to the field’s stratigraphic orientation, and partly to an excess of caution. Since extinct creatures cannot be directly observed, the project of interpreting their structure in functional terms struck many researchers as impossibly speculative. Sure, it is possible to provide functional interpretations of fossils, but with no way of validating these interpretations it was difficult to know what to make of them; and this left the whole enterprise mired in a disreputable kind of uncertainty.
Enter Martin Rudwick. Before embarking on a wildly successful career as a historian of geology, Rudwick was a brilliant young paleontologist with a penchant for brachiopods. These are superficially clam-like invertebrates, which are not, in fact, closely related to clams. (They are not even mollusks.) Brachiopod paleontologists in the 1940s and ‘50s spent much of their time describing new specimens, constructing evolutionary trees—basically, cataloging the diversity of the group. However, what interested Rudwick was not primarily the evolutionary tree of brachiopods. Instead, he was interested in how they lived and behaved as actual organisms, constrained to carve out a living on the seafloor or just above it.

A brachiopod with pronounced zigzag deflections in its valve edges. “Valves” are cupped shells, hinged at the rear, which can be opened for filter feeding
Brachiopods are bit players in the modern marine fauna, but before the end-Permian extinction (ca. 252 Ma) they ranked among the most abundant filter-feeders in the ocean. They were also diverse: far more diverse, in fact, than their living counterparts. So, while it is possible to study living brachiopods as a guide to the anatomy—and to some extent the function—of extinct ones, many forms lack close modern analogs, depriving researchers of a reliable guide for constraining functional interpretations.
This is the predicament that Rudwick’s paradigm method was designed to overcome. If you need a refresher, it goes like this. First, researchers use their knowledge of the principles of engineering to specify an ideal structure, or “paradigm,” that fulfills a postulated function with maximal efficiency under the limitations imposed by the nature of the materials and the evolutionary history of the lineage. Then they conduct tests to evaluate how well the actual structure measures up to the paradigm, or—if there are multiple hypotheses under consideration—paradigms. Rudwick has recently recounted the history of paradigm analysis in a pair of highly readable articles in the Journal of the History of Biology. The second one even cites me of all people, in connection with the subject of this essay. (For those interested in a more detailed primer, Rudwick’s 1964 paper in The British Journal for the Philosophy of Science is a good place to start.)
An example will illustrate how the paradigm method works. Certain fossil brachiopods have valve edges that look a bit like a sawtooth wave. (Valves are cupped shells that press together, as Richard Fortey somewhere says, like hands in a prayer.) The valves must gape apart if food particles are to reach the feeding organ, which scientists know from studies of living brachiopods. But what is the function of the zigzag deflections? Rudwick suspected that the answer had to do with the exclusion of oversized particles during filter feeding. An earlier researcher, Herta Schmidt, had suggested that deflections confer protection relative to an undeflected set-up either by “increasing the area of the slit without increasing the risk of entry of harmful particles, or by reducing that risk without reducing the area” (Rudwick 1964, 136). This suggests a paradigm, or ideal valve edge, in which the zigzag slit “is uniformly narrow at all points, and in which the deleterious effects of the crests (in allowing the passage of particles larger than the rest of the slit) is reduced as much as possible” (137). The matter is complicated by the fact that actual brachiopod commissures are invariably curved; but anyway, the basic design principle ought to hold for curved as well as for straight valve openings.
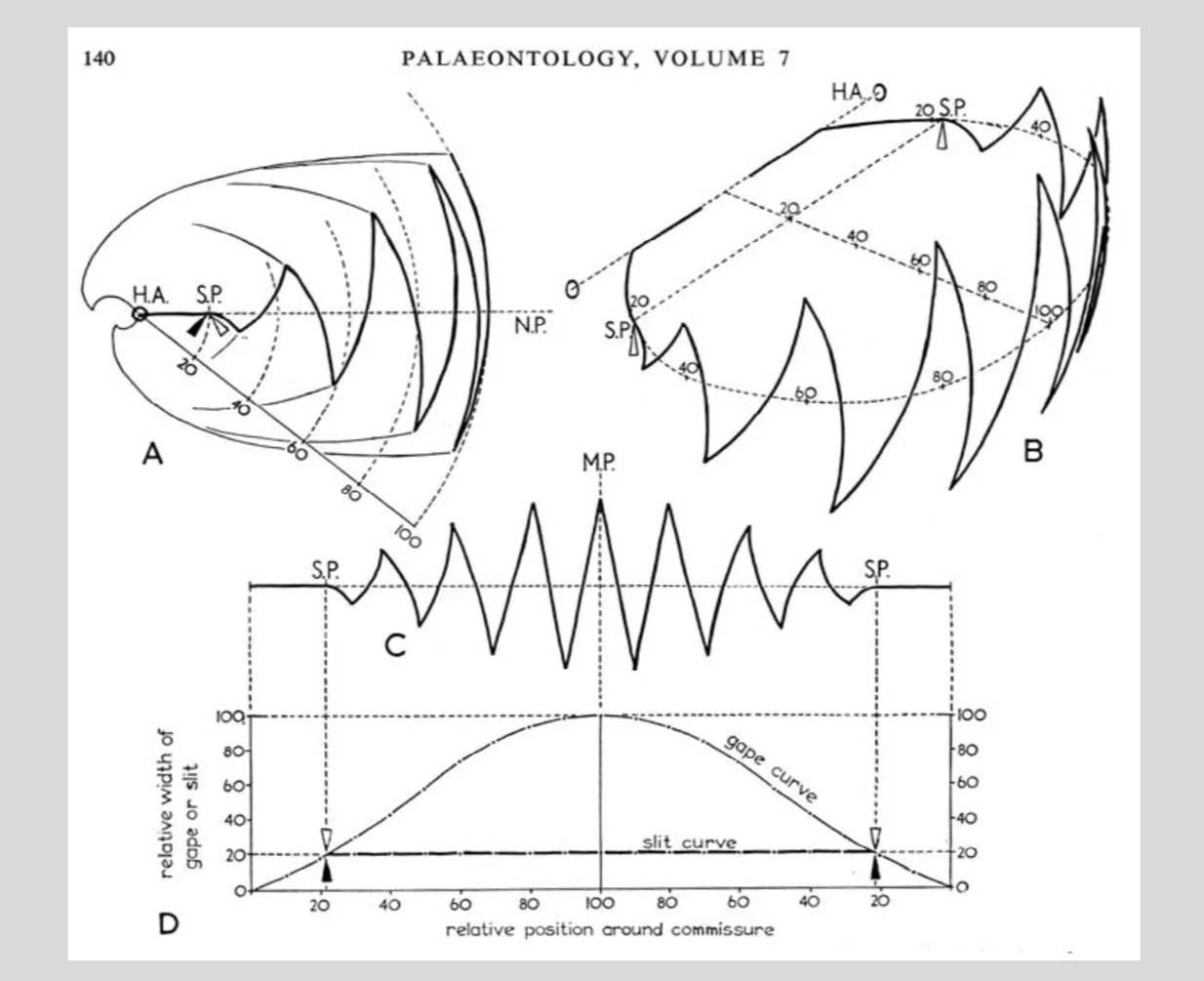
The paradigm for a protective zigzag deflection for a bivalve that is subcircular in plan and biconvex in shape. Here the “wavelength” and “amplitude” of the zigzag are arbitrarily chosen. What matters is that when the valve edges gape apart, this geometrical form will produce a slit of uniform width (except near the hinge and at the crests of the zigzag—see the paradigm for a protective zigzag slit, above). From Rudwick (1964)
Rudwick’s argument is dense, and I am not going to follow out all the complexities here. Suffice it to say that he proceeded to test the function of the zigzag deflections by comparing the shape of actual valve edges to the paradigm structure. And what he found was that “in many cases their valve edges [are] strikingly close to the paradigm, with its strongly arcuate [= curved] form and the distinctive way the ‘amplitude’ of the zigzag [dies] away when traced toward the hinge axis” (Rudwick 2017, 168). According to his later recollection, “[this] proved… that when in life the valves gaped apart through any small angle, there would have been a slit of uniform width between the valve edges all round the shell.” And this in turn permitted him to infer that the zigzag slit was capable of functioning efficiently as a means of excluding harmful particles from the mantle cavity, lending credence to the functional interpretation.

{kind=link}
Drawings of actual brachiopod shells, from Rudwick (1964)
Rudwick made a number of similar studies before leaving paleontology for a full-time gig as a historian of science. But his scientific interests were not limited to the demonstration of adaptation in fossils. He was also interested in the evolution of brachiopods as a whole, including the repeated evolution of structures like zigzag deflections across the brachiopod tree. Rudwick described his “long-term research plan” (retrospectively) as an attempt “to interpret the morphological features of fossil brachiopods of any and every geological period in terms of their likely functions and adaptations, and hence ultimately to reinterpret the large-scale phylogeny of brachiopods in terms of the evolutionary history of the animals’ modes of life” (Rudwick 2017, 148). The results are summarized in a 1970 book, Living and Fossil Brachiopods, based partly on research performed with his student Richard Cowen.
This exercise is basically what Gould had in mind when he spoke about the need for evolutionary paleontologists to “specify biological tasks, define the structures that fit them best and monitor the evolutionary changes that lead, usually in independent lineages, to new grades or functional… zones” (Gould 1970, 111). I was unaware of the extent to which Rudwick had anticipated (and probably inspired) Gould’s vision for “longitudinal paradigm analysis” when I first wrote on this topic back in 2017. As a result I probably overstated Gould’s originality. What Gould brought to the table was not the idea of applying paradigm analysis longitudinally, but rather his emphasis on mechanical improvement as the main vector of history in major taxa. This reflected his Huxleyan inheritance as well as his ambition to find a unifying theme for evolutionary paleontology. Anyway, it was an addition to what he found in Rudwick.
References
Dresow, M. 2017. Before hierarchy: the rise and fall of Stephen Jay Gould’s first macroevolutionary synthesis. History and Philosophy of the Life Sciences 39:1–30. [This was my first paper, and boy does it show. I am grateful for the opportunity to basically rewrite it here]
Dresow, M. 2019. Gould’s laws: a second perspective. Biology & Philosophy, https://doi.org/10.1007/s10539-019-9698-7.
Eldredge, N. and Gould, S.J. 1972. Punctuated equilibria: an alternative to phyletic gradualism. In T.J.M. Schopf (Ed.), Models in Paleobiology, pp. 82–115. San Francisco: Cooper & Co.
Gould, S.J. 1966. Allometry and size in ontogeny and phylogeny. Biological Reviews 41:587–640.
Gould, S.J. 1967. Evolutionary patterns in Pelycosaurian reptiles: a factor-analytic study. Evolution 21:385–401.
Gould, S.J. 1968. Ontogeny and the explanation of form: an allometric analysis. Memoir (The Paleontological Society), Vol. 2, Supplement to Volume 42 of the Journal of Paleontology, pp. 81–91.
Gould, S.J. 1970. Evolutionary paleontology and the science of form. Earth-Science Reviews 6:77–119.
Gould, S.J. 1976. Grades and clades revisited. In R.B. Masterton, W. Hodos and H. Jerison (Eds.), Evolution, Brain, and Behaviour, pp. 115–122. Hillsdale: Lawrence Erlbaum Associates.
Gould, S.J. 2002. The Structure of Evolutionary Theory. Cambridge, MA: Belknap Press.
Huxley, J. 1932. Problems of Relative Growth. New York: The Dial Press.
Rudwick, M.J.S. 1964. The inference of function from structure in fossils. The British Journal for the Philosophy of Science 7:27–40.
Rudwick, M.J.S. 1970. Living and Fossil Brachiopods. London: Hutchinson.
Rudwick, M.J.S. 2017. Functional morphology in paleobiology: origins of the method of “paradigms.” Journal of the History of Biology 50:1–44.
Rudwick, M.J.S. 2018. The fate of the method of “paradigms” in paleobiology. Journal of the History of Biology 51:479–533.
Sepkoski, D. 2012. Rereading the Fossil Record: The Growth of Paleobiology as an Evolutionary Discipline. Chicago: University of Chicago Press.