
* This the latest installment of “Problematica.” It is written by Max Dresow…
The Gaia hypothesis has a Darwin problem. Or at least that is how it has seemed to many scientists loath to accept the idea that self-regulating feedback mechanisms maintain our planet in a state comfortable for life. The main criticism has been that such an arrangement could hardly have arisen by natural selection and is unlikely to have arisen by any means in a world ruled by Darwinian forces. This is because
natural selection operates according to the simple rule of favoring that which works best in the here and now, with no forethought of future implications or… global impact. (Tyrrell 2013, 34)
In other words, nature is unlikely to select for Gaian behaviors and will tend to actively undermine them when they chance to arise. In a world where short-term benefit often clashes with long-term beneficence, tender Gaia hardly stands a chance.
That is the traditional view, anyway, still held by many distinguished scientists. But since about 2014, interest in the possibility of “Darwinizing” Gaia has begun to percolate, stimulated by one of Gaia’s oldest critics. Back in 1981, W. Ford Doolittle declared Gaia “inconsistent with everything we now think we know about the evolutionary process” (Doolittle 1981, 58). Now he thinks that claims like this display a lack of imagination. Gaia is not inconsistent with Darwinism provided we understand natural selection in a sufficiently capacious way. It is even possible that the planet boasts “biosphere-level adaptations, selected by nature for their stability-promoting functions”—the very possibility that critics of Gaia have been most reluctant to countenance.

“Gaia” as seen from space
In a later essay I will explore the question of whether the Gaia hypothesis needs to be “Darwinized.” (To restate the question, I am interested in whether any entity answerable to the description of “Gaia” requires a Darwinian explanation.) For now, let me just say that I am skeptical. I cannot see that “Darwinization” is a suitable aim, at least given what we presently know about the workings of the Earth-life system. But that’s a discussion for another time. Presently, it will suffice to dig into the history of Gaia thinking, which happens to be damn interesting in its own right.
A brief notice before I begin. What follows is a history of the Gaia hypothesis focusing especially on its run-ins with evolutionary theory. It covers some familiar territory, but in covering this territory it avoids a common historiographical pitfall. As Sébastien Dutreuil (2018) points out, a popular notion has it that Gaia sparked a major controversy in evolutionary biology following the publication of Lovelock’s Gaia: A New Look at Life on Earth (1979). This is wrong. Indeed, it is questionable that there was any real controversy at all, since “controversy” implies engagement and the general attitude toward Gaia was one of curt dismissal. Still, advocates of Gaia took the criticisms voiced by evolutionary biologists seriously, as did their opponents. This means that there was a debate about Gaia and evolution; it just happened to be conducted outside of evolutionary biology.
A scientific nonconformist
James Lovelock died on July 26, 2022, exactly 103 years after he was born in Hertfordshire to a family of Quakers. His religious convictions were short-lived, but something of the spirit of nonconformity seems to have been bred in the bone, for Lovelock became the quintessential scientific nonconformist. He is best remembered for developing the Gaia hypothesis, which holds that Earth’s surface is maintained in a habitable state by self-regulating feedback mechanisms involving the biosphere. That is the sober, modern formulation, anyway. In the heady days of the 1970s and 80s, Lovelock often described Earth itself as a living thing—“the largest organism in the Solar System,” as he put it in 1986. An implication was that evolutionary functions could be assigned to certain of Earth’s “organs,” and indeed Lovelock made just this claim in some of his writings. He even argued that Gaia had evolved to maintain surface conditions in an optimal state for life, suggesting that the outcome was more than a byproduct of dynamics whose engine was uncoupled from the interests of the biosphere at large.

An early drawing of Gaia, by James Lovelock. Below it is an electrical circuit for comparison. Source: Dutreuil (2018)
Lovelock came to Gaia through his work at the Jet Propulsion Laboratory, where he was employed as a NASA contractor helping to make scientific instruments spaceworthy. While there he pestered scientists tasked with devising experiments to test for the presence of life on Mars.* To his dismay, he learned that these were based on the assumption that Martian life forms would closely resemble Earthly ones, at least in certain evolved particulars. But this was a risky assumption, since there was no way to know in advance whether Martian organisms would utilize a certain metabolic pathway, say. A more promising approach would be to analyze the Martian atmosphere for signs of chemical disequilibrium, which could betoken biological activity. And as it happened, data on the composition of the Martian atmosphere soon became available. What they revealed was that the atmosphere is chock full of carbon dioxide, along with some nitrogen and noble gasses—exactly what you would expect from a dead planet. But this only highlighted how anomalous Earth’s atmosphere is, with its relatively scant carbon dioxide and abundant molecular oxygen. Why hadn’t more of Earth’s oxygen reacted with carbon to form carbon dioxide? And how could it be that this strange cocktail had remained relatively stable over periods of time much longer than the residence times of constituent gasses? Something must be constantly manipulating the atmosphere from the planet’s surface, the likeliest candidate being life itself.
[* This was 1965, the same year that Mariner 4 returned the first high-quality images of the Martian surface.]
The name “Gaia” was famously suggested by the novelist William Golding, who in addition to being Lovelock’s neighbor was an enthusiast for the idea that the planet is a living thing. It made its debut in (Lovelock 1972), but had a proper coming-out party two years later in a paper co-authored with contrarian microbiologist Lynn Margulis. As Michael Ruse explains, “Lovelock and Margulis staked [their argument] on the dog that barked in the night, or rather on the planet that should have heated up.” The key observation was that Earth’s radiation budget has increased by about 30% over the past three billion years, but the planet has not warmed to a proportional degree. Why? Lovelock and Margulis ventured that life itself has counteracted this warming, partly by altering the composition of the atmosphere. (Hence the title of their paper: “Atmospheric homeostasis by and for the biosphere.”) They also suggested that life may actively regulate surface pH and climate, although these were more tentative suggestions lacking any hint of a mechanism.

Lovelock and Margulis at Lovelock’s home in Devon, 1983. Behind them is the Earth goddess herself
This work failed to attract much attention despite some flashy claims, like the claim that the atmosphere “appears to be a contrivance specifically constituted for a set of purposes.” What really put Gaia on the map—and in the crosshairs—was an engaging little book published in 1979. This was Lovelock’s Gaia: A New Look at Life on Earth, which set out a more expansive version of the hypothesis than had previously been articulated. Lovelock claimed to have written the book “mostly to stimulate or entertain,” and there is no reason to doubt the sincerity of these remarks. Still, his ideas met with a chorus of criticism, some (though not all) of it richly warranted. Perhaps the most consistent theme is that Lovelock’s Gaia is problematically teleological. Lovelock speculated, for instance, that algal mats may have evolved the ability to change color as a means of regulating global temperature. (By turning black, these mats would have lowered the reflectance of Earth’s surface, permitting the conservation of heat.) This seemed to imply that mats evolved in order to perform this function: it was because algal mats were able to regulate global temperature that they (or their capacity to darken in response to cooling) evolved in the first place. But this arguably put the cart in front of the horse, while ignoring a more plausible explanation of the capacity. Other suggestions were even wackier, like the notion that the Great Barrier Reef may be a “partly finished project for an evaporation lagoon” (Lovelock 1979, 91) or the idea that “large mammals” now “serve mainly to provide [certain ancient bacteria] with their anaerobic environment” (102). Perhaps these were the parts of the book intended merely to entertain. Anyway, they opened the hypothesis up to a hailstorm of criticism.
Lovelock contra Darwin
Early criticisms of Gaia tended to sort into two types. The first concerned matters of engineering and focused on the mechanisms of Gaian self-regulation. How does Gaia “know” that temperatures are dropping too low, or that the sea is becoming too salty, such that she can mobilize a countervailing response? And how exactly are these responses implemented by the biosphere? On such matters Lovelock had relatively little to say, but he did have a general rejoinder. That is, where no mechanisms were presently known, we could at least infer from an array of facts that some mechanism(s) must exist. So the absence of evidence of a mechanism does not provide evidence of Gaia’s absence.
The second criticism was harder to brush off and concerned whether Gaia was compatible with evolutionary theory. As Ford Doolittle observed:
It is certainly correct to say that no serious student of evolution would suggest that natural selection could favor the development in one species of a behavior pattern which is beneficial to another… if this behavior were either detrimental or of no selective value to the species itself. (Doolittle 1981, 60–61)
But the Gaia hypothesis seemed to require many species to perform actions that benefit other species at a cost to themselves: and this ought to make Gaian species susceptible to freeriders. Compounding the problem, the benefits of Gaian behaviors are very remote, making it difficult to see how they could ever be targeted by natural selection:
It is difficult to accept that behaviors whose effects... will not be felt for thousands of generations can be selected for, especially when the first beneficiaries of those effects may be organisms which are not themselves responsible for them. The construction of an evaporation lagoon for the sequestration of sea salt may benefit the biosphere as a whole, in the very long run, but what does it do for the organisms who construct it, especially in the short run? (Doolittle 1981, 61)
Richard Dawkins expressed a similar concern in The Extended Phenotype:
I very much doubt that [natural selection for Gaian behaviors] could be made to work… For instance, if plants are supposed to make oxygen for the good of the biosphere, imagine a mutant plant which saved itself the costs of oxygen manufacture. Obviously, it would outreproduce its more public-spirited colleagues, and genes for public-spiritedness would disappear. It is no use protesting that oxygen manufacture need not have costs: if it did not have costs, the most parsimonious explanation of oxygen production in plants would be the one the scientific world accepts anyway, that oxygen is a by-product of something the plants do for their own selfish good” (Dawkins 1982, 236).
Basically, it strained credulity to think that Gaian behaviors could evolve by standard, individual-level selection. But if these behaviors did not evolve by natural selection, how exactly did they evolve?

W. Ford Doolittle, a major player in both the early and the latest stages of the Gaia story
One possibility was that the emergence of Gaia involved higher-level selection—selection for improved planetary homeostasis, say. But it is difficult to see how this proposal could be made to work. For Gaia to have evolved by planetary selection,
[the] Universe would have to be full of dead planets whose homeostatic regulation systems had failed, with, dotted around, a handful of successful, well-regulated planets of which Earth is one. [However, even] this improbable scenario is not sufficient to lead to the evolution of planetary adaptations of the kind Lovelock proposes. In addition we would have to postulate some kind of reproduction, whereby successful planets spawned copies of their life forms on new planets. (Dawkins 1982, 236)
This is perhaps not quite as absurd as Dawkins implies. Meteors sometimes travel between planets and might plausibly enclose a raft of hardy microorganisms. If these were able to seed life on a planet, Gaia-like entities could be said to reproduce by a kind of interplanetary propagation. But the scenario can hardly be called likely. Anyway, the deeper point is a conceptual one. Natural selection involves differential reproduction and evolution by natural selection involves the hereditary transmission of selected traits. Neither is obviously present in the case of Gaia, so planetary natural selection lacks a causal engine.
For many biologists, these criticisms provided sufficient grounds for rejecting all talk of Gaia (Ruse 2013). I have already cited Doolittle and Dawkins. In addition, the latter’s nemesis, Stephen Jay Gould, called Gaia “a metaphor, not a mechanism,” with the implication that there was no Gaian mechanism to be found (Gould 1988). John Maynard Smith upped the ante, labeling the whole Gaia phenomenon an “evil religion” (at least according to Lovelock’s later recollections). And not to be outdone, population ecologist Robert May branded Lovelock a “holy fool.” You get the idea. Gaia was as roundly rejected as any idea submitted to the scrutiny of evolutionary biologists since early models of group selection. The only polite thing for the Earth Goddess to do would have been to roll over and die.
Gaia lives?
Of course, to roll over and die is the very antithesis of what Gaia is about, and would have been anathema to James Lovelock. So Gaia went on, with help and encouragement from influential scientists like Rene Dubos and Philip Morrison. It scored a major victory in 1985 when a week-long conference on whether Earth is a “living organism” was held in Amherst, Massachusetts. This was followed by a second and more wide-ranging conference three years later, funded by the American Geophysical Union. (The first conference was sponsored by the Audubon Society of all things.)
At the second conference, “specialists presented and debated the pros and cons of [the] Gaia Hypothesis from diverse perspectives: modern and ancient biology, ecology, biochemistry, the physicochemical systems of the Earth, oceans, and atmosphere, and the evolution of the solar system” (Kauffman 1988). An especially important paper was given by the geomorphologist James Kirchner, which later appeared as Kirchner (1989). According to Kirchner, the Gaia hypothesis is not one hypothesis but several. These can be arranged in an ascending series beginning with the mundane claim that life and its environment mutually influence one another (“Coevolutionary Gaia”) and culminating in the claim that the biosphere shapes the environment in a way that is optimal for life (“Optimizing Gaia”). In between is a more interesting alternative, which states that life influences the environment via negative feedback mechanisms to maintain Earth in a habitable state (“Homeostatic Gaia”). Actually, Kirschner identified two versions of Homeostatic Gaia: a weak one, which says that the dominant interactions between life and its environment are stabilizing, and a strong one, which says that these interactions make Earth’s surface “significantly more stable” than they would otherwise be. Much subsequent discussion has concerned whether the weak or strong version of Homeostatic Gaia can be maintained, since everyone accepts Coevolutionary Gaia and almost no one can stomach Optimizing Gaia.
At this point I will drop any pretense of writing a linear history of the Gaia hypothesis and will instead focus on a key strand of research. Back in 1983, Lovelock teamed with a former doctoral student, Andrew Watson, to build a heavily idealized simulation model. The idea was to show that planetary self-regulation could arise by a purely mechanical process, thereby defeating the objection that Gaia was problematically teleological. The model imagines a planet orbiting an increasingly luminous star, like our Sun. On the planet lives a population of daisies. The daisies come in two varieties: white ones that reflect radiant light, and black ones that absorb it. At first the planet is too cold to support either variant, but as radiant energy increases black daisies proliferate, warming the planet to the point where white daisies begin to spread.* Further warming favors the proliferation of the white variants at the expense of black ones, with the effect that more radiation is reflected and global temperature begins to drop. At this point black variants begin to spread again. Eventually, the two populations reach an equilibrium, which stabilizes temperature at an intermediate value. This goes to show that the temperature of “Daisyworld” can be stabilized through a wholly blind process, and one that casts the “biosphere” in a leading role.
[* Black and white daisies exhibit the same growth response to temperature, however black forms are assumed to be 10° C warmer than white ones in equivalent ambient conditions and so approach their optimal growth-temperature when ambient temperatures are cooler.]
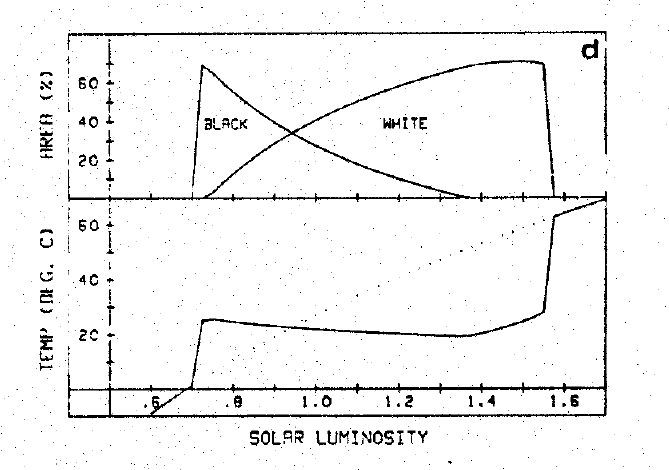
The thermostatic behavior of “Daisyworld.” On the top is a graph of the area covered by black and white daisies, respectively, as solar luminosity increases. Below it is a graph of temperature, which exhibits stability over a wide range of forcings. From Watson and Lovelock (1983)
As Watson and Lovelock observed, “Extrapolation from daisyworld to the earth is, to say the least, rather tenuous” (Watson and Lovelock 1983, 288). Daisyworld does not have so much as an atmosphere to disturb the petals of its inhabitants. It is an exceedingly humble place. Still, its creators thought that it pointed to a kind of system that might exist on Earth. They ask the reader to suppose that the net effect of life on Earth is to reduce atmospheric carbon dioxide and also that the biota is temperature limited. In this case, “a decrease in temperature would lead to an extension of the barren polar regions and would decrease the average level of biological activity over the earth as a whole, while a temperature increase would have the opposite effect. But a decrease in biological activity would presumably also decrease those activities which tend to reduce atmospheric [carbon dioxide]. Thus carbon dioxide would increase to oppose the original change.” They conclude that this represents “a [rudimentary] temperature stabilization system for the earth analogous to that on [D]aisyworld” and one that “may have played a part in regulating the temperature and other environmental variables over the long history of the earth.”
Whether or not this is compelling, what is indisputable is that Daisyworld provided a jumping-off point for studies of the emergence and stability of planetary self-regulation (Wood 2008). Not the least important thing it did was inspire many variants and extensions of itself: one- and two-dimensional Daisyworlds, for instance, and Daisyworlds with rabbits to nibble the daisies. But it also provided a framework for answering the Darwinian challenge. The original Daisyworld contained little to placate an exasperated Darwinian, in part because there are no cheats in Daisyworld: just white and black flowers engaged in a zero-sum bid for world domination. So Lovelock built a variant of Daisyworld that included a free-rider—in this case, a gray daisy that produced no pigment and so outgrew its black and white competitors (Lovelock 1992). He then looked to see what influence the cheat would have on the emergence of self-regulation. As Timothy Lenton explains,
The cheat did not destroy regulation because it had a selective advantage only when the solar input was close to the level at which regulation was not required. At extremes of solar input, the cost of producing the right pigment [was] outweighed by the benefit from being at a temperature closer to the optimum for growth. This emphasizes that environmental regulation can only emerge from traits that are more beneficial than costly to the individual. (Lenton 1998, 443)
Hardly a result to shake the foundations of evolutionary theory, you might think—but for Lovelock this was an adequate response to the worries raised by Doolittle and Dawkins.

Timothy Lenton, a former doctoral student of Andrew Watson and leading Gaia theorist
The biggest outstanding problem with the model—at least when viewed as a response to evolutionary criticisms—is the meager amount of evolution it permits (Barlow and Volk 1992). Indeed, in all early versions of Daisyworld, evolution is restricted to changes in the proportion of static types, all of which are present, in some sense, at the beginning of the simulation. To address this concern, Lenton built a variant of Daisyworld that begins with a population of gray daisies and allows mutations to occur in either direction. At the individual level, “Gaian” and “anti-Gaian” changes are equally likely to occur; mutations are phenotypically random. However, some mutations bring organisms closer to their optimal growth-conditions, and these confer an advantage on the mutants relative to the unmodified type(s). What Lenton found was that when the frequency of mutation is high enough to permit the origin of new types, self-regulation reliably emerges (Lenton 1998, 444). But like all Daisyworld simulations, this hinges on the premise that traits affect the local and planetary environment in the same direction and that this affords the sole basis for selection among competing varieties. Probably this is rare, since in the real world many traits with neutral or harmful environmental effects confer a selection benefit on their carrier (Free and Barton 2007). So at most Daisyworld shows that planetary self-regulation can emerge under particular and perhaps unusual conditions.
Lenton published his model in a review article in Nature titled “Gaia and natural selection.” In the same article he provided a conceptual model for reconciling Gaia and natural selection based on a “hierarchy of feedbacks” (see the figure below). At the lowest level of the hierarchy are purely geochemical and geophysical feedbacks, like the carbonate-silicate thermostat.* To these are added “feedbacks on growth,” or traits whose environmental side-effects either amplify or dampen population growth rates. Such traits provide a basis for environmental regulation, but since they affect carriers and non-carriers in the same way they can provide no selective advantage to carriers. Finally, at the top of the hierarchy, are “selective feedbacks,” or traits whose spread alters the force of selection acting on those traits via some environmental effect. Steps up the hierarchy are often additive, Lenton implies, so a planet with selective feedbacks layered atop non-selective ones is likely to self-regulate better than a planet with just non-selective ones. In this way, the refurbished Gaia hypothesis sought to “bridge the spatial and temporal gaps between the operation of natural selection and the mechanisms of planetary regulation” (Lenton 1998, 439).
[* The carbonate-silicate cycle is sometimes described as a “thermostat” because rates of silicate rock weathering are sensitive to temperature. When temperatures are warm, increased rainfall leads to increased silicate rock weathering, which removes carbon dioxide from the atmosphere and causes temperatures to drop. However, carbon dioxide is ultimately returned to the atmosphere when seafloor carbonates combine with silicate oxide in the mantle, producing carbon dioxide that is vented into the atmosphere through volcanoes.]
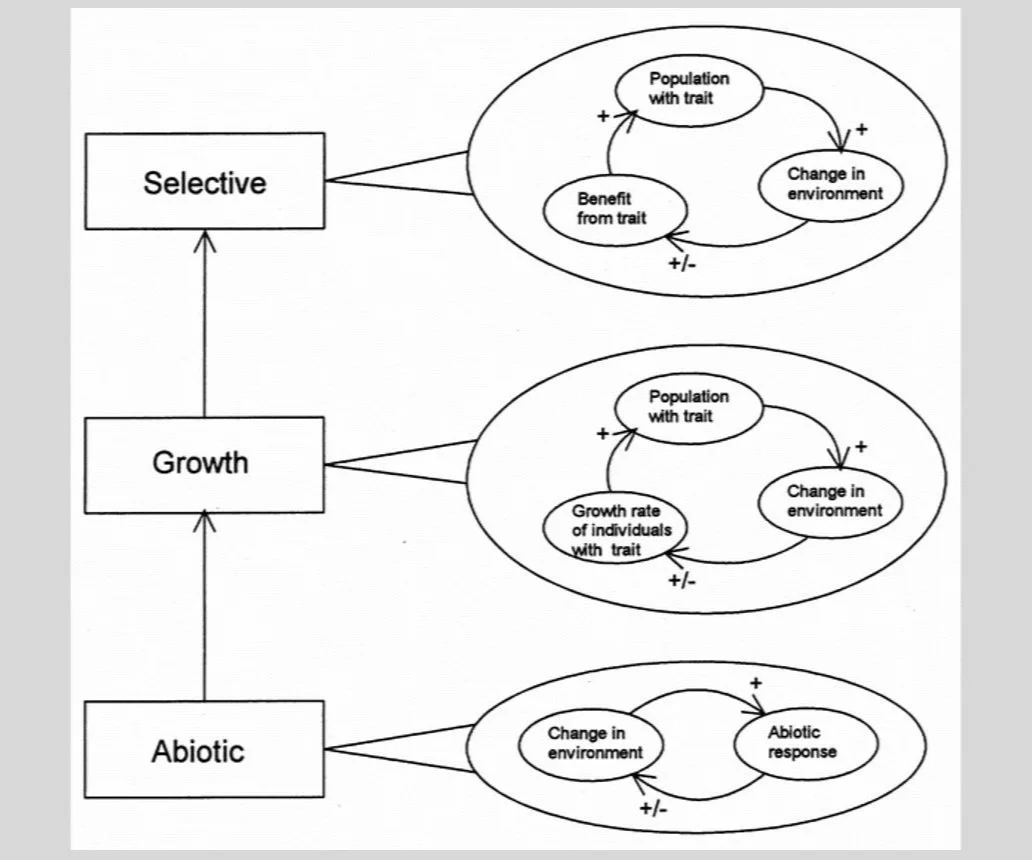
Lenton’s hierarchy of feedbacks, beginning with purely abiotic ones and proceeding through “feedbacks on growth” to “selective feedbacks.” Both feedbacks on growth and selective feedbacks involve traits that produce environmental side-effects. However, in the case of feedbacks on growth, this side-effect influences the growth of trait carriers and other organisms equally, whereas in selective feedback the side-effect confers a special advantage on the carrier.
Yet the most significant part of this article, as far as I’m concerned, is not the conceptual model itself. It is rather the challenge that motivates it, which in Lenton’s words is “to explain how Gaian properties could arise from selection at lower levels” (Lenton 1998, 440). Put more explicitly, the challenge is to show how planetary self-regulation can emerge from selection operating on individuals, sometimes, at least, favoring traits in virtue of their environment-altering properties. This amounts to a recognition that Doolittle and Dawkins had identified a key stumbling block for the Gaia hypothesis. Gaia would have to clear the Darwinian hurdle or die trying. It was a deal with the devil in the details.
FROM DAISYWORLD TO THE WASTELAND
Lenton’s article was not the only publication to address Gaia and natural selection in 1998. In addition, the ecologist Tyler Volk brought out a book, Gaia’s Body, in which he argued that Gaia itself did not evolve at all. By this he meant that selection for Gaian behaviors probably played no role in the emergence of global-scale environmental regulation. Instead, the relevant feedbacks were assembled from the by-products of traits selected for other reasons, and so require no special explanation. (Lenton and David Wilkinson also observed that by-products are an important part of the Gaian story, but it was Volk who promoted the term “by-product” and so achieved a fruitful reframing of the issue)
To understand this emphasis by-products, it is useful to rewind the tape. Back in 1988, a graduate student named Ken Caldeira attended the second Gaia conference (as did Volk, who would later serve on Caldeira’s dissertation committee). While there he became interested in what became known as the “CLAW hypothesis,” which states that a feedback loop including dimethylsulfide (DMS) functions to regulate Earth’s reflectivity, and so its climate. The hypothesis is named after its originators, Charlson, Lovelock, Andreae and Warren, who described the idea in a paper of 1987:
The major source of cloud condensation nuclei (CCN) over the oceans appears to be dimethylsulfide, which is produced by planktonic algae in sea water and oxidises in the atmosphere to form sulfate aerosol. Because the reflectance (albedo) of clouds (and thus the earth’s radiation budget) is sensitive to CCN density, biological regulation of the climate is possible through the effects of temperature and sunlight on phytoplankton populations and dimethylsulfide production. (Charlson et al. 1987, 655)
Put differently, the CLAW hypothesis holds that marine plankton, when pushed to an extreme of their coping range (too hot and sunny or too cold and dim) respond by altering DMS emissions to either increase or decrease solar input to the ocean surface. This in turn will drive the system towards the middle of the plankton’s coping range, opposing the change in temperature (Ayers and Cainey 2007).

Clouds over the southern Pacific Ocean as seen from the International Space Station
Caldeira wondered whether this capacity to alter climate could have evolved by natural selection. He thus set out to determine whether any possible fitness benefit could offset the metabolic costs incurred by the plankton in producing DMSP (the chemical precursor of DMS). He found that they could not, by a long shot. Even using conservative estimates of cost and liberal ones of benefit, the former still outweighed the latter by a factor of a billion or more. This led Caldeira to conclude that “neither climate modulation nor altruism could have been the primary factors in the evolution of mid-ocean DMS production” (Caldeira 1989, 732). While a feedback system involving DMS may serve to regulate temperature, phytoplankton most likely produce DMSP for reasons having to do with cellular osmoregulation, not because of any direct selection on environmental feedback. This means any DMS leaked into the ocean is best interpreted as a happy accident.
But does this mean that the CLAW hypothesis is incompatible with natural selection? Not at all, Volk (1998) answers. It just means that phytoplankton do not produce DMS in virtue of its effects on climate. These confer too paltry and diffuse a benefit to offset the costs of production, so mid-ocean DMS cannot be explained by direct selection for climate modulation. Still, DMS is no less a world-altering chemical for being a “no-cost by-[product] of local… organismic selection” (Volk 1998, 439). Perhaps it is even the case that most Gaian complexity is woven from waste products, making “Gaia” a kind of recycling system comparable to an autocatalytic metabolism (Bagley and Farmer 1992; Lenton and Watson 2011). If this is the case, then our atmosphere is less a biological contrivance than a garbage dump, and something that presumably requires no selective explanation.
Welcome to the wasteland.
* * *
I will conclude this essay by distinguishing two claims about how Gaia came into being and the role that natural selection played in that process. The first is that global-scale self-regulation evolved by natural selection for Gaian behaviors; in other words, that the emergence of Gaia involved selection acting on traits that (1) produced large-scale environmental effects and (2) cost their producers something to produce. This was the position that got Lovelock into so much trouble in the 1980s. It was also the view implicit in the Daisyworld model, and in the many variants of this model that appeared in later decades.
The second view is that global-scale self-regulation emerged through a process that involved natural selection, but that did not include selection for Gaian behaviors. This is the idea that life’s effects on the global environment are predominantly by-products of natural selection operating in local populations. Gaia, on this view, is “one big wasteworld” whose inhabitants exercise great influence over surface conditions without tightly regulating them (Volk 2003, 243). Gaian complexity is “woven for free from excreted material by-products”; it is not assembled by natural selection for the good of the system.
I should say that these views do not have to be framed as mutually exclusive alternatives. If Gaia is a wasteworld, it might still be a wasteworld in which traits occasionally spread in virtue of their “life-enhancing effects” (Lenton and Wilkinson 2003). This requires that these effects be more lavishly enjoyed by a trait’s possessors than by free-riders; but while this is probably rare in nature, it is not impossible. Anyway, the question is on some level an empirical one. The point I wish to emphasize in closing is that the task of “Darwinizing” Gaia looks rather different depending on whether you regard Gaia as a glorified garbage dump or a sprawling organism. I will take up this issue more explicitly in a later essay.
References
Ayers, G.P., and Cainey, J.M. 2007. The CLAW hypothesis: a review of the major developments. Environmental Chemistry 4:366–374.
Bagley, R.J., and Farmer, J.D. 1992. Spontaneous emergence of a metabolism. In Artificial Life II, ed. C. Langton, C. Taylor, J.D. Farmer, and S. Rasmussen. Redwood City: Addison-Wesley.
Barlow, C., and Volk, T. 1992. Gaia and evolutionary biology. BioScience 42:686–693.
Caldeira, K. 1989. Evolutionary pressures on planktonic production of atmospheric sulphur. Nature 337:732–734.
Charlson, C., Lovelock, J.E., Andreae, M., and Warren, S. 1987. Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate. Nature 326:655–661.
Dawkins, R. 1982. The Extended Phenotype: The Gene as the Unit of Selection. Oxford: Oxford University Press.
Doolittle, W.F. 1981. Is Nature motherly? The Coevolution Quarterly 29:58–63.
Doolittle, W.F. Is the Earth an organism? 2020. Aeon. https://aeon.co/essays/the-gaia-hypothesis-reimagined-by-one-of-its-key-sceptics.
Dutreuil, S. 2018. James Lovelock’s Gaia hypothesis: “A New Look at Life on Earth” . . . for the life and the earth sciences. In Dreamers, Visionaries, and Revolutionaries in the Life Sciences, ed. O. Harman and M.R. Dietrich, 272–287. Chicago: University of Chicago Press.
Free, A., and Barton, N. 2007. Do evolution and ecology need the Gaia hypothesis? Trends in Ecology and Evolution 22:611–619.
Gould, S.J. 1988. Kropokin was no crackpot. Natural History 97:12–21.
Kauffman, E.G. 1988. The Gaia controversy: AGU'S Chapman Conference. Eos 69:763–764.
Kirchner, J.W. 1989. The Gaia hypothesis: can it be tested? Review of Geophysics 27:223–235.
Kirchner, J.W. 2002. The Gaia hypothesis: fact, theory, and wishful thinking. Climatic Change 52:391–408.
Lenton, T.M. 1998. Gaia and natural selection. Nature 394:439– 447.
Lenton, T.M., and Wilkinson, D.M. 2003. Developing the Gaia theory: a response to the criticisms of Kirchner and Volk. Climatic Change 58:1–12.
Lovelock, J.E. 1972. Gaia as seen through the atmosphere. Atmospheric Environment 6:579–580.
Lovelock, J.E. 1979. Gaia: A New Look at Life on Earth. Oxford: Oxford University Press,
Lovelock, J.E. 1986. Gaia: the world as a living organism. The New Scientist 18:25–28.
Lovelock, J.E. 1992. A numerical model for biodiversity, Philosophical Transactions of the Royal Society B 338:383–391.
Lovelock, J.E., and Margulis, L. 1974. Atmospheric homeostatis for and by the biosphere: the Gaia Hypothesis. Tellus 26A:2–10.
Ruse, M. 2013. Gaia: Science on a Pagan Planet. Chicago: University of Chicago Press.
Tyrrell, T. 2013. On Gaia: A Critical Investigation of the Relationship between Life and Earth. Princeton: Princeton University Press.
Volk, T. 1998. Gaia’s Body: Toward a Physiology of Earth. Cambridge (MA): The MIT Press.
Volk, T. 2003. Gaia is life in a wasteworld of by-products. In Scientists Debate Gaia: The Next Century, ed. S.H. Schneider, J.R. Miller, E. Crist, and P.J. Boston, 27–36. Cambridge (MA): The MIT Press.
Watson, A.J., and Lovelock, J.E. 1983. Biological homeostasis of the global environment: The parable of Daisyworld. Tellus 35B:284–289.
Wood, A.J., Ackland, G.J., Dyke, J,G., Williamns, H.T.P., and Lenton, T.M. 2008. Daisyworld: a review. Review of Geophysics 46:https://doi.org/10.1029/2006RG000217.
More on Gaia
Lord knows this post is already long enough, but I wanted to draw your attention to a number of interesting videos.
Here is one (actually a series of videos) featuring James Lovelock talking about the science of Gaia.
Here is Lynn Margulis speaking before a crowd of NASA scientists in the 1980s.
And here is Bruce Clarke and Sébastien Dutreuil discussing their new book, Writing Gaia, which chronicles the Lovelock-Margulis correspondence (the book is wonderful too!).